


 Per le sue caratteristiche di alcol-tolleranza e per la generale purezza fermentativa, la specie d’elezione per la conduzione della fermentazione alcolica in enologia è S. cerevisiae. Tuttavia, in passato sono state proposte altre specie di lievito, recentemente riconsiderate per alcuni caratteri tecnologici e sensoriali. Tra le specie non-Saccharomyces più interessanti per la loro influenza positiva sul profilo aromatico dei vini possiamo citare Candida zemplinina, Hanseniaspora uvarum, Metschnikowia pulcherrima, Toluraspora delbrueckii e Zygosaccharomyces bailii. Tali specie sono perlopiù proposte per fermentazioni in cocoltura o in successione scalare con S. cerevisiae, per compensare le eventuali difficoltà a completare la fermentazione.
Per le sue caratteristiche di alcol-tolleranza e per la generale purezza fermentativa, la specie d’elezione per la conduzione della fermentazione alcolica in enologia è S. cerevisiae. Tuttavia, in passato sono state proposte altre specie di lievito, recentemente riconsiderate per alcuni caratteri tecnologici e sensoriali. Tra le specie non-Saccharomyces più interessanti per la loro influenza positiva sul profilo aromatico dei vini possiamo citare Candida zemplinina, Hanseniaspora uvarum, Metschnikowia pulcherrima, Toluraspora delbrueckii e Zygosaccharomyces bailii. Tali specie sono perlopiù proposte per fermentazioni in cocoltura o in successione scalare con S. cerevisiae, per compensare le eventuali difficoltà a completare la fermentazione.
Perché migliorare geneticamente i lieviti?
La scelta del ceppo di lievito ad uso enologico è il fattore che maggiormente influenza il successo della fermentazione alcolica dei vini, per cui è necessario che il ceppo impiegato possegga le caratteristiche o i tratti desiderati. In natura è difficile trovare i ceppi dotati della combinazione di tutti i tratti desiderati, perciò è necessario attuare programmi di selezione e miglioramento genetico. Tali strategie si basano su analisi genetiche e screening funzionali puntuali, che esulano dalla mera provenienza geografica dei lieviti. Nel caso della vinificazione, i criteri di selezione sono stati inizialmente definiti da Giudici e Zambonelli nel 1992, poi ripresi e attualizzati da numerosi Autori (Head, Rainieri e Pretorious, Giudici). La continua rivisitazione dei criteri di selezione, ovvero dei tratti enologici richiesti ad un buon ceppo di lievito enologico, è dovuta al fatto che la lista dei tratti desiderati è aperta ed in continuo divenire, perché alcuni tratti perdono di importanza e nuove caratteristiche vengono richieste, sulla scia delle innovazioni tecnologiche e dei desiderata dei consumatori.
Le collezioni microbiche
Un qualsiasi progetto di miglioramento genetico non può prescindere dalla disponibilità dell’adeguato materiale biologico, che nel caso dei lieviti enologici, è reperibile isolando direttamente da vini a fine fermentazione e ottenuti senza l’aggiunta volontaria di lieviti selezionati. Infatti, le condizioni estreme del mosto a fine fermentazione esercitano una forte pressione selettiva, operando di fatto una preselezione a favore dei ceppi con caratteristiche tecnologiche interessanti, quali la buona capacità fermentativa, la tolleranza all’alcol e, se usata, la resistenza alla solforosa. Le operazioni di campionamento di ceppi da mosti in fermentazione, di caratterizzazione genetico-funzionale e di screening vanno sotto il nome di selezione clonale. La selezione clonale non solo rappresenta il punto di partenza per la realizzazione di nuovi starter enologici, ma soprattutto consente di costituire il pool di materiale biologico (e di caratteri ad esso associati) su cui operare la progettazione di nuovi ceppi ricombinanti attraverso strategie di miglioramento genetico.
Le collezioni microbiche, o biobanche, mettono a disposizione competenze e servizi non solo per la semplice conservazione dei ceppi isolati e selezionati, ma anche per l’archiviazione, con specifiche banche dati informatizzate, delle informazioni ad essi correlati: più informazioni associate ai rispettivi ceppi sono disponibili, maggiore è la qualità e l’utilità della collezione. In particolare, i programmi di miglioramento genetico necessitano di informazioni di tipo genomico, trascrittomico, proteomico e metabolomico, perché consentono di prevedere e/o programmare il risultato finale, attraverso il ricorso a tecniche computazionali di systems biology.
Gli approcci di systems biology
L’elevato numero e la complessità genetica di molti caratteri riducono la possibilità di riscontrare in un unico ceppo la combinazione dei determinanti genetici e dei rispettivi caratteri fenotipici desiderati, per cui è necessario ricorrere a tecniche di miglioramento genetico per ottenere i ceppi evoluti in grado di esprimere i caratteri d’interesse.
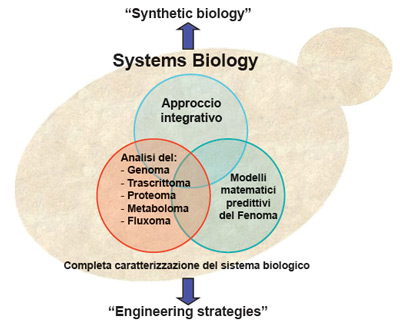
Diverse tecniche di miglioramento dei lieviti sono state ideate e applicate con successo, tra cui: induzione di mutazioni, ibridizzazione, rare-mating, fusione di sferoplasti, over-expression e/o delezione di geni esistenti e introduzione ex-novo di geni eterologhi mediante clonazione e ricombinazione omologa. L’ingegneria metabolica si fonda su solide conoscenze delle vie biochimiche, oggetto della manipolazione genica, e la fase analitica – ossia la conoscenza delle attività cellulari – precede la fase sintetica, ovvero la modificazione genetica vera e propria, che avviene attraverso approcci diretti e mirati quali modificazioni del tipo o della lunghezza dei promoter genici, delezioni o inserzioni di nuovi geni coinvolti nel metabolismo d’interesse.
Questi approcci trovano largo impiego in biotrasformazioni industriali, ad esempio per la produzione di metaboliti o farmaci, ma non sono impiegati in ambito enologico per diverse ragioni, fra le quali il rifiuto da parte dei consumatori ad accettare prodotti ottenuti con organismi geneticamente modificati (OGM).
S. cerevisiae è da tempo usato come modello per lo studio e la descrizione quali/quantitativa dei processi cellulari: è facile da coltivare e da manipolare geneticamente, ha un genoma relativamente piccolo (6000 geni privi di introni) ed è stato la prima cellula eurcariotica il cui genoma è stato completamente sequenziato. Tutto ciò fa si che le informazioni su S. cerevisiae siano moltissime e disponibili in diverse banche dati fra le quali, Saccharomyces Genome Database (SGD-http://www.yeastgenome.org) e Yeastnet (yeast metabolic network - http://yeast.sourceforge.net). Questa disponibilità di informazioni, nonché la sua efficienza come biocatalizzatore di numerosi processi fermentativi, ha determinato la scelta di S. cerevisiae come organismo modello per gli approcci di systems biology.
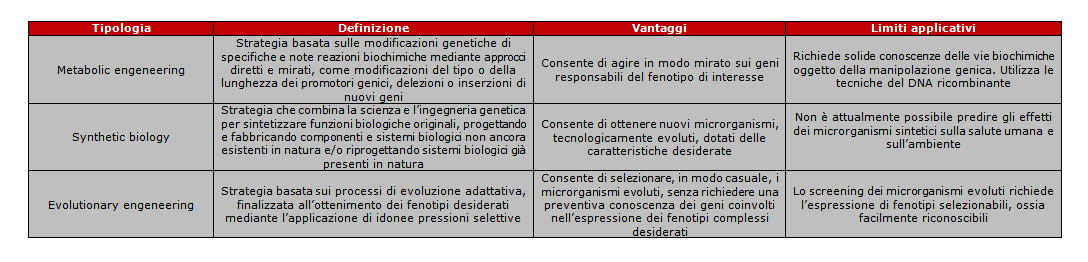
Le strategie di ingegneria genetica vanno intese nella loro accezione ampia ed includono la metabolic engineering, la synthetic biology e la evolutionary engineering; una sintesi sul significato dei termini è riportato in tabella.
Evoluzione adattativa e QTL
La maggior parte dei caratteri, fra i quali anche quelli prettamente tecnologici come il vigore fermentativo, la resa e la tolleranza all’etanolo o la temperatura di crescita, variano in maniera quantitativa nella popolazione e dipendono da più loci (Quantitative Trait Loci o QTL), distribuiti nel genoma e ciascuno dei quali responsabile solo di una piccola parte della proprietà fenotipica quantitativa. Ad esempio, è stato osservato che in S. cerevisiae il carattere tolleranza all’etanolo coinvolge più di 250 geni. Nonostante i recenti successi nel mappaggio dei QTL, grazie all’impiego di nuove tecniche, come l’analisi dei microsatelliti o del polimorfismo di singoli nucleotidi (SNPs), l’identificazione delle basi genetiche di caratteri complessi è considerata a tutt’oggi come uno dei traguardi più ardui della ricerca genetica. Nel caso dei QTL il numero delle variabili genetiche in gioco è così elevato e le interazioni fra queste così complesse e difficili da predire che la costituzione di ceppi geneticamente migliorati è più facilmente praticabile attraverso approcci combinatoriali ciechi, capaci di agire a livello dell’intero genoma.
Le strategie di evolutionary engineering o evoluzione adattativa si prestano al miglioramento di QTL sensibili a una eventuale pressione selettiva. In breve, un organismo è soggetto a una coltura seriale o continua per diverse generazioni, in condizioni non ottimali di crescita (sotto pressione selettiva), al fine di selezionare i ceppi evoluti esprimenti il fenotipo d’interesse. L'evoluzione adattativa privilegia le mutazioni che risultano vantaggiose per la cellula sottoposta a condizioni di stress. Le mutazioni, per definizione, determinano la formazione di nuovi alleli nella popolazione e forniscono la variabilità genetica su cui agisce la selezione. Pertanto, l’evoluzione adattativa determina l'aumento nella popolazione delle cellule varianti che più si adattano all'ambiente caratterizzato dalla pressione selettiva. La procedura è efficace, ma poiché è basata sull’occorrenza occasionale di mutazioni durante la riproduzione mitotica può richiedere tempi lunghi.
L’evoluzione adattativa, di fatto, è il risultato della selezione di fenotipi selezionabili, cioè sensibili alla pressione selettiva esercitata dal mezzo e dalle condizioni di crescita. Caratteristiche quali la resistenza alla SO2, l’alcol tolleranza o la capacità di sviluppare in presenza di elevate pressioni osmotiche sono legate a ricombinanti direttamente selezionabili. Tuttavia molte altre caratteristiche di interesse enologico tra cui, ad esempio, la produzione di composti sensorialmente favorevoli, sono legate a variazioni e ricombinazioni genetiche che non esprimono fenotipi direttamente selezionabili, su cui non è possibile operare approcci di evoluzione adattativa.
La mimica della selezione naturale
Un altro limite intrinseco degli approcci di evoluzione adattativa è la loro applicabilità a fenotipi selezionabili, cioè sensibili alla pressione selettiva esercitata dal mezzo e dalle condizioni di crescita. Approcci di evoluzione adattativa non sono applicabili a caratteristiche di interesse enologico tra cui, ad esempio, la produzione di composti sensorialmente attivi, che sono legate a variazioni e ricombinazioni genetiche che non esprimono fenotipi direttamente selezionabili.
In laboratorio è possibile attuare pressioni selettive indirette anche su fenotipi non selezionabili, nel caso si riescano ad individuare uno o più step, nella pathway biosintetica del metabolita di interesse, sottoponibili a pressione selettiva.
Un esempio concreto è riportato di seguito e riguarda la non produzione di solfiti e solfuri e l’elevata produzione di glutatione, sfruttando la capacità dei ceppi evoluti di svilupparsi in presenza di metalli pesanti, quali cromo e molibdato, molto tossici per le cellule nella forma ionica esavalente.

Metalli pesanti come fattore di pressione selettiva
La resistenza al cromato e al molibdato costituisce lo step selezionabile per l’individuazione dei ceppi caratterizzati da un alterato metabolismo dei solfati. In particolare, la resistenza delle cellule ad alte concentrazione di ioni Cr6+ e Mo6+ può essere dovuta ad almeno due meccanismi: 1) una mutazione a carico dei geni responsabili della codifica delle proteine trasportatrici degli ioni Cr6+ e Mo6+, le stesse proteine di membrana che mediano l’entrata del solfato; 2) un aumento nella sintesi del glutatione, un tiolo costituito da cisteina, glicina e acido glutammico, che svolge un’importante azione antiossidante ed è in grado di chelare i metalli pesanti formando complessi che vengono trasferiti attivamente nel vacuolo tramite trasportatori presenti nella membrana del vacuolo stesso. Nel vacuolo i complessi metallo-glutatione vengono degradati dagli enzimi litici e gli amminoacidi e i metalli liberati vengono immagazzinati in questo organello per poi essere riutilizzati o eliminati dalla cellula.
La riduzione dei solfati a solfiti e solfuri sono step necessari della sintesi degli aminoacidi solforati cisteina e metionina.
La degradazione di quest’ultimi determina la produzione del glutatione. In sintesi, abbiamo osservato che i nuovi ricombinanti ottenuti, resistenti al molibdato e al cromato, producevano quantità bassissime di solfiti e solfuri, oppure producevano quantità elevate di glutatione, importante antiossidante naturale (vedere approfondimento sul web).
La strategia descritta ci ha consentito di ottenere un ceppo con buonissima attitudine enologica, basso produttore di solfiti e solfuri e non producente tioli sensorialmente sgradevoli.
[box title= "Un esempio di collezione microbica" color= "#c00"]
La Food and Industrial Microbial (FIM) collection del Dipartimento di Scienze della Vita dell’Università di Modena e Reggio Emilia dispone di circa 2.700 ceppi, di cui circa 1.600 ceppi di lievito isolati da diverse matrici alimentari ma soprattutto da mosti e vini di diversa provenienza. Tutti i ceppi in collezione sono sottoposti ad accurata caratterizzazione fenotipica e molecolare e le informazioni sono elaborate con il software GelCompar II (Applied Maths), che consente di normalizzare e comparare fra loro i dati di fingerprinting raccolti su ceppi isolati da matrici differenti.[/box]
[box title= "Potenzialità degli OGM" color= "#c00"]
L’uso di OGM in enologia non è consentito, per cui le engineering strategy che impiegano tecniche del DNA ricombinante sono qui riportate più per dovere di cronaca che per un attuale impiego enologico. Va tuttavia ricordato che esperienze di laboratorio hanno ampiamente dimostrato l’utilità delle tecnologie del DNA ricombinante per il miglioramento dei lieviti enologici. Ad esempio sono stati costituiti ceppi più efficienti nell’utilizzo degli zuccheri, con capacità di chiarificazione dei vini, con aumentata attività peptolitica, glucanolitica o xilanolitica e influenti sull’aroma del prodotto finale per l’espressione dell’attività β-(1,4)-endoglucanasica.[/box]
[box title= "Evoluzione adattativa e lieviti autoctoni" color= "#c00"]
In apparenza i principi che regolano l’evoluzione adattativa, in primis la pressione selettiva esercitata dall’ambiente ‒ nel nostro caso dal sistema vigneto-cantina-mosto sulla popolazione dei lieviti ‒ supporta la tesi, tanto cara a molti Autori, della presenza dei lieviti autoctoni. Quei ceppi di lievito che nel tempo si sono evoluti e selezionati in un determinato territorio, ceppi che sono in grado di valorizzare al meglio le note di tipicità del vino del territorio di riferimento. In realtà quanto esposto in questo articolo sulla evoluzione adattativa confuta in modo documentato e concreto tale convinzione. Infatti, l’evoluzione adattativa funziona per i caratteri selezionabili e quindi sensibili alla pressione selettiva esercitata dall’ambiente circostante. La domanda che viene spontanea è: qual è la pressione selettiva che privilegia i ceppi che migliorano la tipicità territoriale dei rispettivi vini di riferimento? Non c’è alcuna pressione selettiva, ma se talora capita di isolare un ceppo in grado di avere tante caratteristiche desiderate ed anche quella di essere eccellente per la fermentazione dei vini del proprio territorio, è solo perché il caso è un burlone e si prende gioco di noi presentandoci anche gli eventi meno probabili.[/box]
Articolo a firma di
Luciana De Vero, Francesco Mezzetti, Lisa Solieri, Paolo Giudici
Università degli Studi di Modena e Reggio Emilia
La ricerca qui presentata è stata svolta con il contributo di AEB (Brescia)
Approfondimenti a cura degli Autori
PER APPROFONDIRE
Come cambiano i criteri di selezione dei lieviti
Nell’enologia poco tecnologica del passato, l’uso di alte dosi di solforosa era pratica necessaria per l’ottenimento di vini accettabili e, di conseguenza, ai ceppi di lievito era richiesta la proprietà di tollerare elevate concentrazione di solfiti. Oggigiorno, il miglioramento significativo delle condizioni igieniche delle cantine ed il maggior controllo della temperatura di fermentazione, del potenziale di ossidoriduzione e di altri parametri di processo, rendono possibile una riduzione dell’impiego della solforosa e conseguentemente l’alta resistenza a questo biocida non rappresenta più un carattere primario di selezione. Al contrario, stanno acquisendo un crescente interesse caratteri funzionali che determinano positive proprietà sensoriali nei vini, come l’espressione di geni codificanti per idrolasi capaci di idrolizzare i terpeni glicosilati o di pathway metaboliche coinvolte nella produzione di tioli sensorialmente positivi. Ne consegue che i programmi di miglioramento genetico per i lieviti enologici sono prevalentemente orientati sui tratti sensorialmente attivi.
Collezioni microbiche: come devono essere gestite
L’importanza delle collezioni microbiche è intrinsecamente connessa all’adeguata caratterizzazione dei microrganismi presenti e all’accuratezza delle informazioni loro associate, motivo per cui la gestione delle collezioni è affidata a curatori esperti e competenti in diverse aree della biologia microbica. In particolare sono necessarie competenze multidisciplinari che spaziano dai processi di isolamento, identificazione e selezione dei ceppi sulla base di proprietà tecnologiche importanti per specifiche applicazioni, alla definizione dei caratteri di potenziale interesse e alla realizzazione delle strategie di selezione e miglioramento genetico per i caratteri desiderati. Solo le collezioni in grado di fornire servizi e ricerca ad alti livelli riescono a soddisfare le aspettative del mercato, riuscendo nell’intento di selezionare i ceppi adeguati per ogni specifica richiesta. Non va poi trascurata la necessità di conservare il materiale biologico preservandone la stabilità genetica, perché durante i processi moltiplicativi delle cellule si verificano mutazioni che nel tempo possono modificare significativamente le caratteristiche delle colture. Per questa ed altre ragioni, i ceppi selezionati vengono conservati in condizioni che ne impediscono la moltiplicazione, come la liofilizzazione ed il congelamento a bassissime temperature-80°Co, ancora meglio, in azoto liquido a-196°C.
Alcune collezioni microbiche sono molto specifiche e si occupano di pochi generi o di soli funghi o batteri, altre più grandi e complesse spaziano dagli organismi cellulari, ai virus, ma anche ai genomi, geni e sequenze di DNA di specifico interesse, che possono costituire materiale biologico di riferimento e/o utile per gli studi di systems biology.
Evoluzione adattativa e ciclo vitale di S. cerevisiae
Il particolare ciclo riproduttivo di S. cerevisiae - si può moltiplicare per gemmazione (divisione solo mitotica) o per via sessuale (meiosi e mitosi) - può essere sfruttato per aumentare considerevolmente il numero di ricombinanti da sottoporre a pressione selettiva. In genere, i ceppi selvaggi presentano ambedue le modalità di moltiplicazione e sono generalmente omotallici, cioè sono in grado di ripristinare lo stato diploide, dopo una divisione meiotica, autonomamente con un processo di riconversione del tipo sessuale tra cellula madre e figlia.
Le mutazioni possono avvenire sia durante la divisione mitotica sia durante quella meiotica, generando individui diversi dal genitore. Inoltre, durante la meiosi, si hanno ricombinazioni fra coppie di cromosomi omologhi, determinando spore, originate da un ceppo selvaggio ed eterozigote, che saranno tutte diverse una dall’altra. Pertanto, sia prima che dopo il ripristino dello stato diploide, tutta la progenie derivata sarà costituita da individui diversi tra di loro. Rispetto a cicli ripetuti di divisioni mitotiche, durante il processo di moltiplicazione sessuale viene amplificata la variabilità genetica del pool di discendenti su cui effettuare la selezione dei ceppi evoluti attraverso l’applicazione di un’idonea pressione selettiva. In termini genetici rigorosi, l’aumento di variabilità in una popolazione è dovuto all’introduzione di nuovi alleli, attraverso le mutazioni, mentre durante i processi di ricombinazione omologa e/o crossing-over non si ha un aumento di variabilità, in senso stretto, ma solo ricombinazione tra cromosomi omologhi. Ciò premesso, va sottolineato che per i QTL - la maggior parte dei caratteri lo sono - non è tanto importante la presenza di nuovi alleli, ma la combinazione, fra quelli esistenti, che esprime il fenotipo ai massimi livelli. Proprio durante la riproduzione sessuale, la ricombinazione è quantitativamente molto rilevante.
Il ruolo del glutatione
In S. cerevisiae, il glutatione è pari a circa l’1% del peso secco cellulare e rappresenta più del 90% dei tioli a basso peso molecolare. Il glutatione viene inizialmente consumato dal lievito nel corso della fermentazione alcolica per essere rilasciato successivamente in seguito alla lisi cellulare. Il glutatione ridotto svolge numerose funzioni nel mosto e nel vino; ben nota è la sua capacità di ridurre gli o-chinoni prodotti dall’azione della polifenolo ossidasi sugli acidi idrossi-cinnamil tartarici, evitando, in questo modo, la formazione di polimeri fenolici responsabili del colore bruno di mosti e vini bianchi. L’elevata produzione di glutatione è importante nei programmi di selezione e miglioramento genetico dei lieviti per uso enologico, in quanto i ceppi con queste caratteristiche possono rappresentare un’alternativa all’impiego dell’anidride solforosa, limitatamente al suo ruolo antiossidante.
Bibliografia
Brown C.J., Todd K.M., Rosenzweig. R.F. (1998). Multiple duplications of yeast hexose transport genes in response to selection in a glucose-limited environment. Mol. Biol. Evol., 15: 931-942.
Ciani M., Comitini F., Manazzu I. and Domizio P. (2010). Controlled mixed culture fermentation: a new perspective on the use of non-Saccharomyces yeasts in winemaking. Fems Yeasts Research, 10: 123-133.
De Vero L., Solieri L. & Giudici P. (2011). Evolution-based strategy to generate non-genetically modified organisms Saccharomyces cerevisiae strains impaired in sulfate assimilation pathway. Letters in applied microbiology 53, 572-575.
Ferea T.L., Botstein D., Brown P.O., Rosenzweig R.F. (1999). Systematic changes in gene expression patterns following adaptive evolution in yeast. Proc. Natl. Acad. Sci. (U.S.A), 96: 9721-9726.
Giudici P., Rainieri S., Pulvirenti A., Zambonelli C. (2006). Strategie di selezione e miglioramento di lieviti di interesse enologico. Industrie delle bevande XXXV: 22-30.
Giudici P., Solieri L., Pulvirenti A., Cassanelli S. (2005). Strategies and perspectives for genetic improvement of wine yeasts. Applied microbiology and biotechnology 66: 622-628.
Giudici P., Zambonelli C. (1992). Criteri di selezione dei lieviti per enologia. Vignevini 9: 9-34.
Head G.M. (1999). Novel yeasts in winemaking-looking to the future. Food Aust. 51: 347-352.
Lavigne, V., Pons, A. & Dubourdieu, D. Assay of glutathione in must and wines using capillary electrophoresis and laser-induced fluorescence detection. Changes in concentration in dry white wines during alcoholic fermentation and aging. Journal of chromatography. A 1139: 130-135 (2007).
Marullo P., Bely M., Masneuf-Pomarede I., Aigle M. & Dubourdieu D. (2004). Inheritable nature of enological quantitative traits is demonstrated by meiotic segregation of industrial wine yeast strains. FEMS yeast research 4: 711-719.
Mendoza-Cózatl D., Loza-Tavera H., Hernandez-Navarro A., Moreno-Sanchez R. (2005). Sulfur assimilation and glutathione metabolism under cadmium stress in yeasts, protists and plants. FEMS Microbiology Reviews, 29: 653-671.
Osteergard S., Olsson L., Nielsen J. (2000). Metabolic engineering of Saccharomyces cerevisiae. Microbiol. Mol. Biol. R. 64: 34-50.
Pretorius, I. S. (2000). Tailoring wine yeast for the new millennium: novel approaches to the ancient art of winemaking. Yeast 16: 675-729.
Rainieri S. & Pretorius I (2000). Selection and improvement of wine yeasts. Annals of microbiology 50: 15-31.
Risch NJ. (2000). Searching for genetic determinants in the new millennium. Nature 405: 847-856.Rosi I., Vinella M. and Domizio P. (1994), Characterization of β-glucosidase activity in yeasts of oenological origin. Journal of Applied Microbiology 77:519-527.
Salgues et al., 1986. Oxidation of grape juice 2-S-glutathionyl caffeoyl tartaric acid by Botrytis cinerea laccase
and characterization of a new substance: 2,5-di-S-glutathionyl caffeoyl tartaric acid. Journal of Food Science 51: 1191-1194.
Stephanopoulos G., Aristodou A., Nielsen J. (1998). Metabolic engineering. Academic, San Diego
Warner J. R., Patnaik R. & Gill R. T (2009). Genomics enabled approaches in strain engineering. Curr Opin Microbiol. 12(3): 223-30.







{kind=link}